
Trilobita
Trilobieten behoren tot de geleedpotigen (Arthropoden) en leefden van het Cambrium tot in het Perm in het Paleozoicum. De groep heeft de massa-extinctie aan het einde van het Perm niet overleefd. De klasse Trilobita is in negen verschillende orden onderverdeeld. De orde Phacopida is misschien wel de bekendste orde van deze negen. Deze negen orden zijn weer onder te verdelen in 150 families met in totaal zo'n 15.000 beschreven soorten. Met enige regelmaat worden nog nieuwe (sub)soorten ontdekt. De trilobieten kunnen een zeer succesvolle groep genoemd worden en vertoonden een hoge vormendiversiteit.
Gerastos catervus, uit het midden-devoon van België (© F Lerouge)
Morfologie
De naam 'trilobiet' is ontleend aan het feit dat het pantser uit drie delen of 'lobben' is opgebouwd, twee pleurale lobben en in het midden een axiale lob. Ook longitudinaal kent een trilobiet drie delen; het cephalon (kopschild), de thorax (lijf, opgebouwd uit segmenten) en het pygidium (staartschild). Het pantser van trilobieten is grotendeels opgebouwd uit calciumcarbonaat, waardoor het relatief vaak bewaard blijft. Dit pantser is heel hard, waardoor de trilobiet zoals de meeste arthropoden tijdens de groeifase verschillende malen moet vervellen. Ook deze vervellingsresten worden fossiel aangetroffen, en vormen wellicht het grootste aandeel van gevonden trilobietfossielen. De voornaamste morfologische elementen van een trilobiet zijn in onderstaande figuur opgenomen.
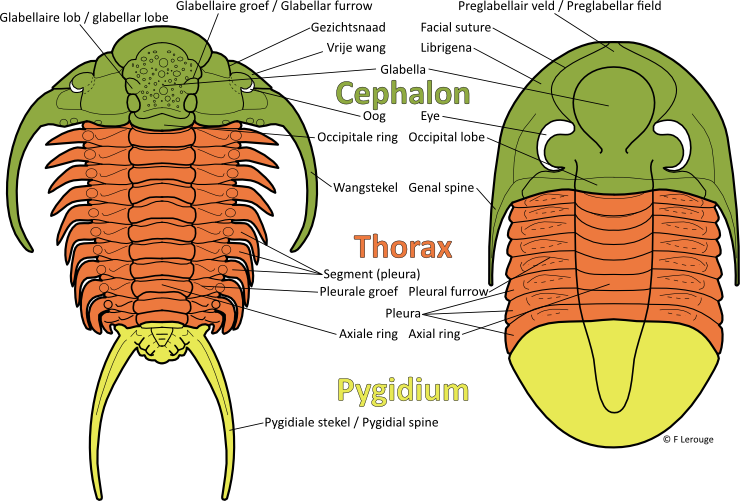
Dorsale morfologie van trilobieten: voornaamste onderdelen
De vorm van trilobieten is erg uiteenlopend, wat indicatief is voor een breed gamma aan levenswijze. Sommige soorten vertonen spectaculaire vormen, met indrukwekkende stekels. Ook andere vormen van ornamentatie ('prosopon') op het pantser zijn gedocumenteerd, waarbij stekels, tuberkels, porieën en terraslijnen zeer frequent voorkomen. De functie van deze elementen is uiteraard zeer uiteenlopend:
- stekels: de meest voor de hand liggende functie van stekels is defensie.
- tuberkels: kan uiteenlopende functies vervullen, van defensie tot versteviging van het pantser, of verhogen of verlagen van de wrijvingsweerstand van het panster, ...
- poriën: komen zeer vaak voor en speelden wellicht een rol bij respiratie, zintuiglijke waarneming of secretie. Sommige poriën penetreren het carapce volledig.
- terraslijnen: een interessante en veel voorkomende vorm van prosopon. terraslijnen verhogen de wrijvingsweerstand in 1 bepaalde richting en wordt vaak aangetroffen aan de onderzijde en randen van het exoskelet. Het wordt aangenomen dat dit het dier hielp om niet weg te zakken in of zich af te zetten tegen fijn sediment.

Dit exemplaar -met schaalbewaring- toont een goed ontwikkelde tuberkels op de glabella (coll. FL).
De ogen van trilobieten zijn gefacetteerd. Er zijn verschillende types gekend:
- Holochroaal: ogen bestaan uit vele kleine enkelvoudige en dicht op elkaar gepakte lenzen.
- Schizochroaal: oog heeft een beperkt aantal dubbele lenzen, van elkaar gescheiden door sclera.
- Abathochroaal: Dit type is beperkt tot een relatief kleine cambrische groep.
De ogen zijn in sommige gevallen zeer goed bewaard gebleven en konden erg gesofisticeerd zijn. Dit gaat in het bijzonder op voor schizochroale ogen, een opmerkelijk type dat binnen de suborde Phacopina is ontwikkeld. Verschillende ogen hebben een tweede complex gevormde lens, waarmee het zicht sterk verbeterd werd doordat uiteenlopende vertekeningen gecorrigeerd worden.


Voorbeelden van een schizochroaal oog (links) en holochroaal oog (rechts) (coll. FL).
De ventrale morfologie van trilobieten is goed gekend, mede dank zijn de vondst van uitstekend bewaarde specimens uit verschillende conservat-lagerstätten. Bij deze vondsten zijn de ledematen en kieuwen, sensorische voelsprieten en soms zelfs sporen van het digestief systeem bewaard gebleven. Dergelijke vondsten kennen we onder meer van de Burgess shale in Canada, de Hunsrück schiefer in Duitsland, de Beecher beds in de USA, Chengjiang in China en de Fezouata shales in Marokko.
Elk segment droeg een paar van dubbele ledematen, telkens een looppoot met een kieuwaanhangsel. Het cephalon, en insommige gevallen ook het pygidium, droeg een paar flexibele antennes, die duidelijk een sensorische functie hadden. Onder het cephalon bevond zich een reeks poten waarvan de basale delen zich rond de mondopening bevonden en deel uitmaakten van het kauwapparaat. Dit was op zijn beurt bedekt met een mondplaat of hypostoom, dat net als het pantser sterk verkalkt was en wel eens fossiel wordt aangetroffen, soms geassocieerd met de triobiet.
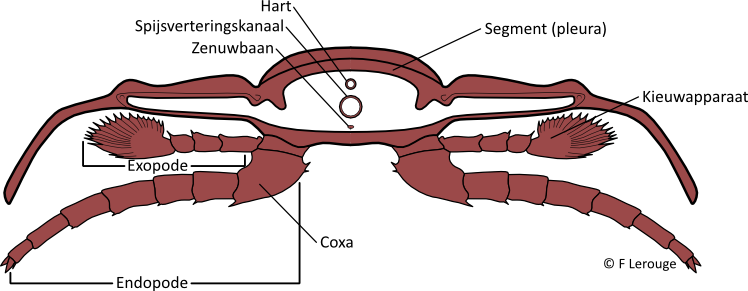
Doorsnede van de thorax
Levenswijze
Trilobieten waren exclusief mariene dieren. In grootte varieerden trilobieten van enkele millimeters tot meer dan een halve meter. Van de tot nu toe grootst gevonden soort, Isotelus rex, is een exemplaar uit New Foundland gekend van 72 cm. Ze hebben doorheen de evolutionaire geschiedenis een breed gamma aan ecologische niches bevolkt, en zowel benthische, demersale als pelagische levenswijzen zijn gerapporteerd. Trilobieten leefden van plankton tot grotere zeedieren. Sommige soorten waren actieve jagers en hadden een goed zichtvermogen. Andere soorten leefden op grotere diepten en hebben het gezichtsvermogen secundair verloren.
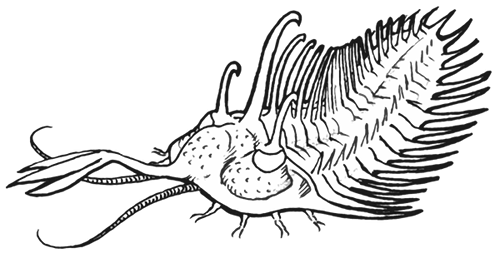
Artist impression van Walliserops, die een opmerkelijke gevorkte stekel droeg waarvan de functie op heden nog een raadsel blijft (© F Lerouge).
Verschillende trilobieten konden zich oprollen. Algemeen wordt gedacht dat dit voornamelijk een verdedigingesmechanisme is geweest. Alle zachte delen van de trilobiet worden zo immers omsloten door het pantser. Bij sommige genera is een duidelijk slotsysteem beschreven, waarbij uitsteeksels van het pygidium in inkepingen aan de rand van het cephalon pasten, waardoor de trilobiet, eens gesloten, vrijwel onmogelijk te opnenen was door een predator.

Voorbeeld van een opgerolde trilobiet (Reedops) (coll. FL)
Dat trilobieten ook gegeten werden, blijkt duidelijk uit de exemplaren die worden gevonden met al dan niet geheelde bijtsporen. Deze vertonen soms secundaire genezing, wat erop wijst dat de trilobiet de aanval had overleefd. Welke predatoren hiervoor verantwoordelijk waren, is niet steeds even duidelijk. Het ordovicium en devoon zijn gekenmerkt door de opkomst van de Gnathostomata (kaakdieren), waaronder binnen de pantservissen ook enkele toppredatoren. Maar ook andere invertebraten, zeeschorpioenen en trilobieten incluis, hebben mogelijk op trilobieten gejaagd.
Het harde pantser dat door evolutie allerlei vormen kon aannemen in combinatie met de flexibele pleura, welke de trilobiet erg mobiel maakte, was de sleutel tot hun succes. Dit harde panster groeide niet mee en moest regelmatig vervangen worden tijdens de groeifase, net zoals bij moderne arthropoden. Hiervoor brak bij de meeste trilobieten het cephalon open vie de suturen, breuklijnen die de losse wangdelen van de rest van het cephalon scheidden, waardoor de trilobiet naar voren uit het pantser kon kruipen. Een individuele trilobiet vervelde meermaals gedurende een leven, en het zijn bijgevolg vaak deze vervellingsresten die fossiel worden aangetroffen. Ze zijn dan vaak herkenbaar aan het (deels) losgekomen zijn van de verschillende onderdelen, één of twee vrije wangen in het bijzonder. Soms zijn vervellingsresten of exuviae zo talrijk aanwezig, dat ze heelder gesteentelagen lijken te vormen.

Een massa vervellingsresten (coll. FL)

Typische vervellingsrest: de librigena ontbreken, en thoraxelementen zijn lichtjes losgekomen, verschoven of bij elkaar getrokken als het dier het pantser van zich afschudde (coll. FL)
Ook sporenfossielen van trilobieten zijn teruggevonden, waarvan de meest voorkomende als het vormgenus cruziana beschreven zijn. Dit zijn kruipsporen. Ook rustsporen zijn gekend (Rusophycus), net als loopsporen (Diplichnites).


Foto's of locaties voor Trilobita bekijken
Feedback
Doneer
Wij zijn geheel afhankelijk van donaties. Daarom vragen wij onze gebruikers ons te helpen.